Обеспечение метаболизма нейронов рассматривается как главная функция мозговой гемоциркуляции. Ее нарушения вызывают тяжелую патологию, нередко завершающуюся трагическим концом. Поэтому борьба с сосудистыми поражениями мозга является первоочередной задачей, успех решения которой связан с полнотой знаний о механизмах регуляции мозгового кровообращения.
Наличие множества механизмов, направленно изменяющих просвет кровеносных сосудов, признано фундаментальным принципом регуляции транспорта крови. Для утверждения этого представления заметную, а в некоторых случаях доминирующую роль сыграли морфологические исследования субстрата "сосудистых механизмов" мозговой гемоциркуляции. С помощью новых методов были расширены и уточнены сведения об эфферентной и афферентной иннервации магистральных, пиальных и внутримозговых артерий; выяснено значение сосудистых эндокриноцитов и эндотелиоцитов в регуляции вазомоторных реакций.
В итого обобщения материалов оформилось учение о системе управления мозговой гемодинамикой, в которую были включены нейро-мышечный, нейро-паракринный и интимальный или эндотелиозависимый механизмы.
Афферентная и эфферентная иннервация. Нейро-мышечный механизм.
Проблема нервной регуляции мозговой гемодинамики традиционно вращается вокруг вопроса морфологическом субстрате и физиологических механизмах, осуществляющих рецепцию и реализующих вазомоторику. Открытый за последние десять лет ряд ранее неизвестных вазоактивных веществ внес не только новые представления в понимание механизмов регуляции, но и одновременно показал большую сложность организации эфферентных и афферентных систем мозговой гемоциркуляции. Тем не менее основные законы, сформулированные в прошлом относительно афферентной иннервации кровеносных сосудов мозга, сохранили своё значение.
Строение рецепторов определяется типом сосуда и ролью, которую он играет в гемодинамике. В стенках артерии имеются древовидные, кустиковые и клубочковые окончания, локализованные преимущественно в местах дихотомии сосудов на ветви. В артериолах пре.эжхз-0обладают компактные шаровидные и вытянутые клубочки диаметром от 12 до 90 мкм с высокой концентрацией нервных волокон. На капиллярах окончания имеют специальные клетки. Для стенок вен более типичными являются диффузные и сплетеневидные чувствительные аппараты. В распределении рецепторов вдоль сосудистого русла отмечена существенная неравномерность: одни значительной длины сегменты артерии лишены рецепторов, в то время, как другие имеют их высокую концентрацию. Наибольшим числом рецепторов клубочковой формы располагают артерии мягкой оболочки диаметром от 40 до 350 мкм на передней поверхности шейного отдела спинного мозга и основания среднего мозга человека.
Сосуды мозга способны активно изменять свой объем. Очевидно, чувствительные образования артерии, их клубочковые и древовидные формы, являются механорецепторами. Описанные в артериях виллизиева круга осумкованные рецепторы ближе стоят к колбам Краузе и тельцам Гольджи-Маццони. Располагаясь на путях тока крови в вещество мозга, эти рецепторы контролируют температуру, определяют пределы ее нижней и верхней границ. Сосуды мозга особенно чувствительны к метаболитам, на которые они реагируют увеличением емкости. Физиологические исследования доказывают наличие в их стенках хеморецепторов, к которым можно отнести окончания на капиллярах и венах, заключающими кровь, насыщенную продуктами обмена. С рецепторами на стенках интракраниальных артерий связывают происхождение головных болей. Электрофизиологические исследования показали, что таковыми должны быть свободные нервные окончания.
Афферентный аппарат сосудов обладает возможностями собирать и передавать весь объем сведений о состоянии мозговой гемодинамике в первичные (спинальные) и вторичные (бульбарные) сосудодвигательные центры. Из чувствительных узлов IX, X, XI пар черепномозговых нервов иннервируются все экстрацеребральные, особенно, магистральные артерии. Аксоны блуждающего нерва своими коллатералиями заканчиваются на нейронах сосудодвигательного центра. Этот факт объясняет механизм, благодаря которому раздражение центрального конца перерезанного блуждающего нерва вызывает не только расширение мозговых сосудов, но и изменяет системную гемодинамику. Основное значение бульбарной чувствительной иннервации заключается в обеспечении определенного уровня притока крови в магистральные артерии. Внутримозговые артериолы, капилляры и вены снабжены чувствительными окончаниями спинальной природы, с помощью которых контролируется органная гемоциркуляция из сегментных центров спинного мозга.
Координированная реакция сосудистой системы реализуется через эфферентное звено, которое не менее разнообразно качественно, а количественно обладает заметным превосходством над афферентной иннервацией. На сегодняшний день с большой полнотой изучена холин и адренергическая иннервация нервных сосудов головного и спинного мозга.
Концентрация холин- и адренергических сплетений находится в зависимости от способа подачи крови к головному мозгу, от преобладания значения в мозговой гомоциркуляции сосудов или каротидной, или вертебральной системы. При прямой, характерной для человека, крысы, отчасти собаки, подаче крови через каротидную систему ее артерии имеют более высокую концентрацию эфферентных волокон. Среди них особой густотой сплетений выделяется средняя мозговая артерия, снабжающая кровью наиболее обширные области полушарий большого мозга. При доминировании окольного кровотока через вертебральную систему, преимущественное число волокон получают, например, у кролика и морской свинки, позвоночная и основная артерии.
Содержание эффекторных волокон соответствует типу сосуда и его роли в гемодинамике. Наиболее высокую концентрацию и наибольшее абсолютное число аксонов находится на магистральных артериях и артериях виллизиева круга. По мере уменьшения диаметра сосудистых трубок стремительнее падает абсолютное содержание, медленнее и более плавно концентрация.
Имеется прямая зависимость между степенью мышечного вооружения сосуда и концентрацией эффекторных аксонов. У степной черепахи в средней мозговой артерии содержится три слоя гладких мышечных клеток, а у кошки 7-8; концентрация проводников у этих животных составляет 32 и 122 аксона/мм2 соответственно.
Вместе с тем концентрация нервных волокон на стенке сосуда не отражает в полной мере степень его иннервированности. Для оценки нейромышечных взаимодействий большое значение имеет количество на аксоне варикозных расширений, которые являются местами накопления и выделения медиатора на иннервируемый субстрат. К примеру, в венозных образованиях твердой мозговой оболочки млекопитающих концентрация адренергических волокон на 1/3 меньше такого же показателя в средней мозговой артерии, а у ныряющих птиц, даже, несколько больше.
Одним из аргументов против участия нервов в контроле за мозговой гемодинамикой считается их малое число на пиальных и внутримозговых артериях. Между тем этот "недостаток" компенсируется высоким значением концентрации варикозных расширений, что соответствует большей чувствительности пиальных артерий и нервным стимулам.
Значение холинергической иннервации оценивается в настоящее время однозначно: ее аксоны являются источниками AX, который вызывает эндотелиозависимую релаксацию артерий и вен. При большой плотности адренергических нервов их роль регуляции мозговой гемодинамики все еще находится в рамках гипотез. Предполагается, что обилие адренергических нервов поддерживает в мозговых сосудах высокий тонус миоцитов, необходимый для защиты мозга от резких повышений системного кровяного давления.
Представления о нервной регуляции кровеносных сосудов головного и спинного мозга значительно расширились благодаря открытию нейропептидов.
Наибольшее внимание исследователей привлекло вещество P (BP), вазоинтестинальный пептид (ВИП) и нейропептид (НУ). Иммуногистохимическими, радиометрическими и биохимическими методами эти пептиды идентифицированы у многих животных и человека. На мозговых артериях крыс 49% нервных терминалей ВИП, 35% - НУ и 5% - ВР. Согласно общепринятой гипотезе ВИП является принадлежностью холинергических аксонов, а НУ адренергических; ВР – находится в афферентных нервных волокнах и окончаниях с электроноплотными везикулами диаметром 120 нм.
Методом ретроградного транспорта и иммунохимическими исследованиями ВР идентифицировано в протонейронах спинномозговых узлов, ганглиев блуждающего и тройничного нерва и в их волокнах.
Полагают, что ВР является модулятором боли. По данным других исследователе ВР имеется в баро- и хеморецепторах мозговых артериол, где вещество выполняет функции нейротрансмиттера. На изолированных мозговых сосудах показано, что ВР обладает способностью релаксировать гладкие мышечные клетки. Предполагается, что с помощью аксон-рефлекса ВР участвует в вазодилятации.
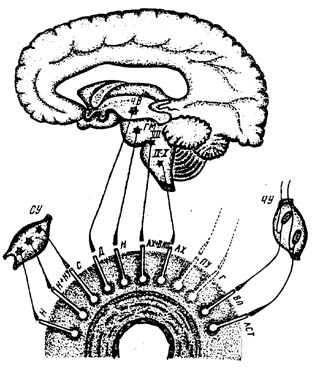
Рис. 1. Нейрохимические типы сосудистых аксонов и их источники.
чу– чувствительные узлы спинномозговые и черепномозговые; су – шейные и верхнегрудной симпатические узлы; чв– черное вещество; гм– голубоватое пятно; VII, IX, X – ядра черепномозговых нервов; н– норадреналин; н+ну– норадреналин + нейропептид V; с– серотонин; д– дофамин; ах+ВИП – ацетилхолин + вазоин-тестинальный пептид; ах– ацетилхолин; пу– пурины; г – гистамин; вп – вещество P; авс –L-аспартат; Темные широкие линии – центры доказаны, черные узкие линии – центры предполагаются, пунктирные линии – центры не известны.
ВИП вызывает независимую от ацетилхолина и следовательно от эндотелия релаксацию интракраниальных артерий и артериол. Пептид заключен в везикулах нервных терминалей, расположенных в адвентиции на расстоянии не менее 100 нм от гладких мышечных клеток. Наличие ВИП-позитивных аксонов в церебральных сосудах человека сомнительно, у крыс их мало, у мышей – умеренно, у кошек обнаружены густые сплетения из нервных волокон, содержащие ВИП. По ходу барабанной струны обнаружена численно ограниченная группа нервных клеток положительно реагирующая на ВИП. Открытие этих нейронов объясняет дилятаторные свойства данного нерва.(Рис. 1.)
Нейропептид У по свойствам является типичным вазоконстриктором; в эксперименте он вызывает уменьшение просвета внутримозговых артериол до 81% их диаметра. Терминали аксона с НУ находятся в наружной оболочке артерии и располагаются на расстоянии 100-120 нм от плазмолеммы гладких мышечных клеток. Предполагается, что НУ является модулятором, повышающим констрикторное действие биогенных моноаминов. Вместе с тем не исключена возможность, что НУ и ВИП могут выполнять функции медиатора, если их содержание в аксоне оказывается преобладающим или исключительным над катехоламинами или АХ.
Помимо холинергических, адренергических, пуринергических и пептидергических в мозговых сосудах допускается наличие аксонов и другой нейрохимической специализации, исследованных пока недостаточно.
Сосудистые эндокриноциты. Нейро-паракриальный механизм.
К клеткам, структурно и функционально связанным с кровеносными сосудами головного и спинного мозга, относятся хромаффиноциты, меланоциты и тканевые базофилы. Обобщенно за наличие в цитоплазме материала, оформленного в виде гранул, их называют гранулоцитами, а за участие в обмене биогенных моноаминов — моноаминоцитами. Все они инкретируют вазоактивные вещества и могут рассматриваться как сосудистые эндокриноциты с паракриновым механизмом действия.
В кровеносных сосудах мозга, помимо типичных тканевых базофилов, описаны нейролипомастоциты, по морфологии и свойствам, занимающие как бы промежуточное положение между истинными гепариноцитами меланоцитами.
Биохимическими и цитофотометрическими исследованиями в тканевых базофилах и хромаффиноцитах показано наличие триптамина, серотонина, гистамина и дофамина. Суммарное содержание индолалкиламинов и катехоламинов у рыб, птиц, млекопитающих животных количественно не различаются, а у человека достоверно снижено. Иммуногистохимическими методами идентифицирован вазопрессин в меланоцитах человека и хромаффиноцитах лягушки, хомяка, крысы, морской свинки, коровы; ВИП и вазопрессин обнаружены в тканевых базофилах крысы.
Располагая биологически активными веществами, эндокриноциты могут функционировать в качестве местной, относительно автономной регуляторной системы, для которой характерно большое морфологическое и функциональное разнообразие и двойственность функциональных отправлений, то есть способность давать положительные и отрицательные эффекты. Кроме того, работа этой системы координинируется нервным механизмом.
Сосудистые эндокриноциты имеют функциональные связи с нервной системой, через которые усиливается или ослабляется их секреторная деятельность.(Рис. 2.)
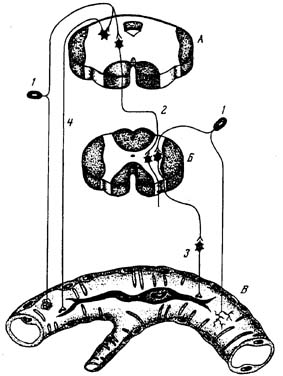
Рис. 2. Регуляция артерии нейропаракринным механизмом.
А – ствол мозга; Б – спинной мозг; В – артерия с сосудистым эндокриноцитом; 1 – протонейрон чувствительного узла; 2 – волокно ретикулоспинального тракта; 3 – постганглионарный адренергический аксон; 4 – преганглионарный холинергический аксон.
Можно считать, что сосудистые эндокриноциты по своему положению занимают место эффекторной клетки, подобное периферическому вегетативному нейрону:
— морфологически и функционально они, как и все эндокринные клетки, связаны с преганглионарными холинергическими аксонами.
— содержат вазоактивные вещества – катехоламины: индолалкиламины, адренолютины, хиноиды и полипептиды.
— при введении экзогенного АХ и других агонистов М-рецепторов реагируют дегрануляцией и секрецией биологически активных веществ;
— антихолинэстеразные вещества, ингибируя АХЭ клетки, вызывают реакцию, сходную при введении АХ;
— реакция клеток полностью подавляется атропином – специфическим блокатором М-рецепторов.
Эти качества сосудистых эндокриноцитов полностью совпадают с рецепторно-секреторными и эффекторными свойствами пара-нейронов.
На сосудистые эндокриноциты помимо холинергических конвергируют адренергические аксоны, терминали которых образуют синаптические контакты с весьма узкой щелью от 7 до 20 нм. Важное значение адренергических связей заключено в том, что их медиатор через b-адренорецепторы тормозит дегрануляцию и секрецию сосудистыми эндокриноцитами вазоактивных веществ. Таким образом, адренергическая реакция иннервация гранулосодержащих клеток выступает как антагонист холинергических связей относительно функций этих клеток.
Интимальный или эндотелиозависимый механизм.
До середины текущего столетия сосудистый эндотелий рассматривался как гомогенный, довольно инертный, клеточный пласт, основное значение которого видели в том, что он разделяет внутри- и внесосудистое пространство, и благодаря ровной поверхности обеспечивает легкость скольжения крови. Всплеск интереса к эндотелию связан с развитием учения о микроциркуляции, что совпало с широким использованием электронной микроскопии и новым методом культивирования сосудистого эндотелия.
В дополнение к таким важным функциям, как транскапиллярный обмен, регуляция липидов плазмы, поддержание гомеостаза, он модулирует реактивность гладких мышц сосудистой стенки:
— предотвращает и определяет уровень активности норадреналина, серотонина, брадикинина;
— превращает предшественники (напр. : ангиотензин I в ангиотензин II; АТФ в аденозин) в сосудистоактивные вещества;
— секретирует сосудистоактивные вещества, например, производные арахидоновой кислоты, главным образом, простациклин;
— освобождает под влиянием АХ внутренний эндотелиальный фактор, релаксирующий гладкие мышцы, - окись азота NO и, секретируя эндотелин, повышает их тонус.
Эти новые концепции о свойствах эндотелия явились завоеванием физиологии последнего десятилетия. Одновременно были представлены функциональные доказательства и морфологический субстрат для реализации эндотелиозависимой (интимальной) регуляции тонуса гладких мышечных клеток. При воздействии АХ на эндотелиоциты, на фоне максимального сокращения аорты кролика под влиянием норадреналина, наступает ее релаксация. Эта работа стала рубежом, с которого начался счет новым фактам о значении эндотелия в регуляции кровеносных сосудов. За время чуть больше 10 лет обследованы практически все крупные артерии и вены, в том числе сосуды головного мозга кролика, крысы, собаки, свиньи, кошки, крупного рогатого скота, человека и низших позвоночных — рыб, амфибий, пресмыкающихся. В 90% опытов зарегистрировано расширение сосуда, наступающее через 6-80 сек. После нанесения АХ на эндотелиальные клетки. Скарификация эндотелия более чем на 40% или обработка его атропином подавляет АХ-зависимое расслабление сосуда. На основании этих данных было сделано два вывода:
— дилятация кровеносного сосуда при аппликации на эндотелиальную выстилку АХ реализуется через мускариновые рецепторы;
— релаксирующий фактор (окись азота NO) синтезируется в эндотелиальных клетках. Для развития реакции необходимо наличие свободного внеклеточного кальция. Добавление в среду Ca2+ионофора А 23187 стимулирует, а его отсутствие или блокада нифедипином кальциевых каналов тормозит релаксацию.
Сравнительно недавно установлено, что окись азота (NO) синтезируется некоторыми нейронами головного мозга и является нейромедиатором, обеспечивая синаптическую и несинаптическую передачу информацию в ЦНС. NO образуется в нейронах и некоторых других клетках при помощи фермента — синтетазы окиси азота из L-аргинина. Этот фермент относится к группе NADPH-диафораз.
Исследования нейронов, синтезирующих NO, показали, что NO не является самостоятельным нейромедиатором, а ассоциирована с другими нейромедиаторами. Так, в неостриатуме NO синтезируют нейроны, вырабатывающие АХ и нейропептид-Y, а в коре большого мозга — некоторые ГАМК-ергические нейроны. Недавно было обнаружено, что различные повреждающие факторы способны индуцировать выработку NO нейронами, ранее ее не синтезировавшими. Показано также, что нейроны, содержащие NO-синтетазу, оказываются более устойчивыми к разного рода повреждающим воздействиям, чем соседние нервные клетки, не вырабатывающие NO.
Достаточно хорошо обоснованным является предположение об участии нейрооксидергических, то есть синтезирующих NO, нейронов в регуляции гемодинамики в головном мозге. NO приписывают также роль ретроградного синаптического медиатора, однако на этот счет имеются противоречивые данные.
В головном мозге NO может синтезироваться не только нейронами, но и глиоцитами, и эндотелиоцитами кровеносных сосудов, в которых присутствуют другие формы NO-синтетазы.
В полушариях большого мозга NO-синтетаза определяется в весьма малом числе (менее 1%) нейронов. Преимущественно такие нейроны локализованы в областях, связанных с обонятельным и зрительным анализаторами, и в неостриатуме. В коре большого мозга встречаются единичные, в основном веретеновидные нитрооксидергические нейроны во II-III и значительно меньше в V-VI слоях. Продукт реакции находится как в телах нейронов, так и в их отростках.
В сером веществе головного мозга расположены сплетения нервных волокон, содержащих NO-синтетазу. В неокортексе выделяют два таких сплетения: поверхностное и глубокое.
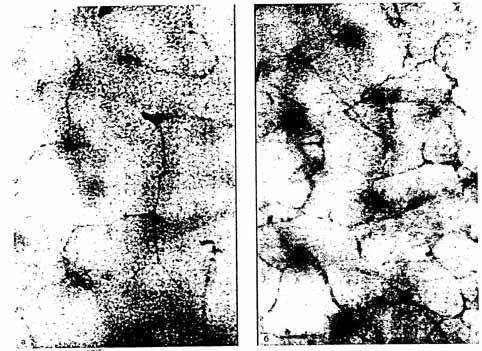
Рис. 3 Сосудистое русло переднего мозга эмбриона человека 10 нед. Развития
а - поверхностный слой; б - субвентрикулярный слой. Тотальный препарат. Реакция выявления НАДФН-диафоразы после фиксации в 4% параформальдегиде.
Отделом морфологии (зав. — проф. В.А. Отеллин) научно-исследовательского института экспериментальной медицины РАМН (Санкт-Петербург) были проведены исследования по выяснению источника NO в неокортексе человека с 7-й по 12-ю неделю эмбрионального развития. Было установлено, что распределение NO-синтетазы вблизи поверхности неокортекса, по всей видимости, связанно с накоплением ее в терминальных расширениях отростков радиальной глии. Отсутствие NO-синтетазы в дифференцирующихся нейронах коры большого мозга указывает на то, что нейроны, вырабатывающие NO, дифференцируются в более поздние сроки развития. NO-синтетаза в нейронах неокортекса человека появляется не ранее 15-й недели эмбрионального развития. (Рис. 3.)
Интенсивная окраска эндотелиоцитов кровеносных сосудов указывает на образование этими клетками сравнительно большого количества NO. Наличие NO-синтетазы в эндотелиоцитах сосудов малого диаметра, что не характерно для взрослых особей (У взрослого человека кровеносные сосуды головного мозга окрашиваются с различной интенсивностью. Наиболее сильная реакция наблюдается в эндотелии крупных артерий, проникающих из оболочек в мозг. По мере уменьшения диаметра артерий количество продукта реакции уменьшается.), может быть связано с тем, что мишенью действия эндотелиальной NO служат не только элементы сосудистой стенки, но и окружающие нейральные элементы. Учитывая тот факт, что эндогенная NO тормозит пролиферацию и стимулирует дифференцировку развивающихся нервных элементов, в то время как развитие нервной системы у мутантных мышей, лишенных гена нейрональной NO-синтетазы, не нарушается, можно полагать, что источником NO, необходимой для развития нервной системы, являются эндотелиоциты кровеносных сосудов.
Существование NO-синтетазы в клетках радиальной глии и распределение фермента вблизи поверхности неокортекса согласуется с представлением о важной роли радиальной глии в регуляции миграции и дифференцировки нейробластов.
Таким образом, полученные результаты свидетельствуют о том, что в период с 7-й по 12-ю неделю эмбрионального развития в развивающейся коре большого мозга человека существуют две популяции клеток, содержащих NO-синтетазу и вырабатывающих NO. Это эндотелиоциты кровеносных сосудов и клетки радиальной глии. По-видимому, глиальные элементы в области барьерных структур мозга могут синтезировать NO и в физиологических условиях, а не только при воспалении. Источником NO в коре и подкорковых ядрах могут служить и собственные нейроны, и нейроны других областей, поскольку NO-синтетаза находится как в перикарионе, так и отростках нейрона. Распределение NO-синтетазы в нейронах свидетельствует о возможности не только постсинаптической, но и пресинаптической ее локализации.
Благодаря обнаружению эндотелиального фактора миорелаксации, установлен механизм возбуждения гладких миоцитов. В момент расслабления гладкой мышцы в ней увеличивается содержание циклического гуанилатмонофосфата (цГМФ). Введение в среду блокатора гуанилатциклазы — метиленового голубого — задерживает релаксацию. Видимо NO активирует зависимую от цГМФ протеинкиназу, которая опосредует расслабление гладких миоцитов (Рис. 4.).
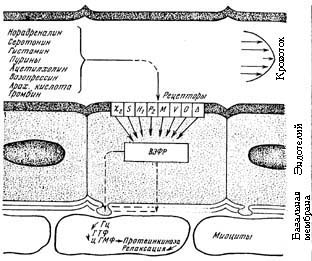
Рис. 4. Механизм эндотелиозависимой релаксации.
Модулирующее влияние эндотелия на гладкие миоциты не только с помощью химических, но и механических факторов передается разными путями, среди которых по общей оценке ведущее значение имеет миоэндотелиальные контакты. Их число увеличивается по мере того, как сокращается калибр артерий и достигает максимальной величины в артериолах, то есть в сосудах, претендующих на исключительное значение в регуляции органной гемодинамики. Симметричные миоэпителиальные контакты образуются за счет базальных выпячиваний цитоплазмы эндотелиоцита и тянущимися на встречу равными по длине выростами гладкого миоцита. Чаще же миоэндотелиальные связи бывают асимметричными за счет большей длины или выпячивания эндотелиальной клетки, или, наоборот, выроста гладкого мышечного волокна. В местах соприкосновения выпячиваний образуется щелевой контакт, через который происходит быстрая и адресная передача сигнала от эндотелиоцита к миоциту. Исследования, выполненные на артериях мозга позвоночных, показали, что у млекопитающих при сравнении с рыбами число базальных выростов увеличивается в 2, их длина в 4, а площадь в 8 раз. Эти количественные данные с миоэндотелиальных контактах находятся в соответствии с возрастающим значением эндотелия у высших животных в реакциях расширения артерий. Помимо миоэндотелиальных контактов, вазоактивные вещества воздействуют на гладкие миоциты из ретроэндотелиального пространства, куда они попадают из крови с помощью пиноцитозного транспорта, осмоса и диффузии.
Интактный эндотелий может быть источником вазоактивных веществ, повышающих тонус гладких миоцитов. Одним из таких агентов является мощный вазоконстрикторный пептид — эндотелин, выделенный из эндотелия микрососудов головного мозга. В состав эндотелина входит 21 аминокислота: он образуется в результате протеолиза из пептида, имеющего около 200 аминокислотных остатков. Эндотелин оказывает выраженное вазоконстрикторное действие на пиальные артерии и вызывает стойкое дозозависимое сокращение средней мозговой артерии, в меньшей степени сокращает просвет пиальных вен.
Эндотелин в значительных количествах высвобождается при гипоксии. В этих условиях страдает функция эндотелиоцитов и вещества, стимулирующие в норме дилятацию, например, тромбин, простациклин и другие, становятся активными вазоконстрикторами. Они усиливают эффекты эндотелина, что может быть причиной спазма артерий мозга.
Многообразие и разнонаправленность реакции сосудов мозга и сердца, опосредуемых через эндотелий, предлагается совокупно выделить в особую систему регуляции функций сердечно-сосудистой системы в связи с фармакологическими возможностями эффективно устранять гемодинамические нарушения.
Заключение
Данные материалы дают основания внести некоторые коррективы в существующие представления о регуляции кровеносных сосудов головного мозга. Прежде всего, под давлением новых фактов теряет свое значение метаболическая теория, так как соображения, положенные в ее основу, нашли объяснения в рамках эндотелиозависимого механизма. Важнейшие метаболиты пуриновых оснований, арахидоновой кислоты, многие другие продукты обмена, циркулирующие в крови, реализуют свои вазоактивные свойства через сосудистый эндотелий. На поверхности эндотелиоцита и в его цитоплазме образуются простогландины, лейкотриены и окись азота NO, вызывающая при морфо-функциональной целостности клеточного пласта релаксацию артерий и вен. Даже такой конечный продукт обмена, как углекислота, которая является главным аргументом адептов метаболической теории, регулирует тонус миоцитов через эндотелиальную выстилку.
Обнаружено, что реакция миоцитов на изменения системного кровяного давления, не прямая, а опосредована эндотелием и зависит от высокой чувствительности его клеток к изменениям скорости кровотока. В ответ на "силу сдвига", то есть величину, на которую меняется скорость кровотока, эндотелиальные клетки с помощью окиси азота NO — эндотелиального фактора миорелаксации — контролируют тонус гладких мышечных клеток.
Многообразие новых данных по нейрохимии и функции интрамуральных аксонов укрепили представления о значении нейрогенного механизма в регуляции сосудов мозга путем прямых и опосредованных через сосудистые эндокриноциты влияний на локомоторные функции гладких мышечных клеток. Наметились связи, устанавливающие пути взаимодействия между регуляциями нейрогенной и зависимой от эндотелия. Это, прежде всего, наличие кооперативности в реализации дилятации между эндотелиоцитами и АХ, который синтезируется в основном холинергическими аксонами. На протяжении сосудистого русла мозга выявлен градиент нервной и интимальной регуляции: по мере сокращения диаметра артерий уменьшается значение нервов и усиливается роль эндотелиальных механизмов; эндотелиозависимая регуляция становится преобладающей для внутримозговых кровеносных сосудов. Представлены материалы о зависимости морфо-функциональной целостности эндотелиального пласта от состояния внутристеночного нервного аппарата артерии.
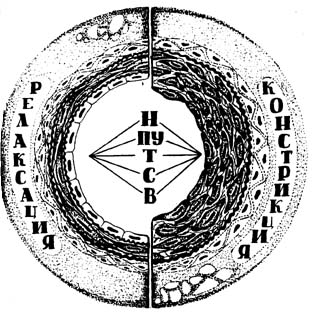
Рис. 5. Схема иллюстрирующая прямое (констрикторное) и опосредованное через эндотелий (релаксирующее) действие норадреналина (н), пуринов (пу), тромбина (т), серотонина (с) и вазопрессина (в) на миоциты артерий.
Мышечную оболочку сосуда можно рассматривать как двухконтурный экран, ограниченный со стороны крови эндотелием, а снаружи — адвентицией с ее нервным аппаратом. Следует допустить, что внутренний контур через эндотелий реализует дилятацию, в то время, как импульсы, действующие из адвентиции на наружный контур вызывают констрикцию. На самом деле, многие вещества обладают свойством понижать или повышать тонус гладких миоцитов в зависимости от того, действуют ли они через эндотелий или прямо на мышечный пласт (рис. 5.). Характерный пример: серотонин, нанесенный на эндотелий стимулирует дилятацию, а приложенный к мышечной оболочке работает, как сильный констриктор, способный вызвать спазм церебральных артерий. Типичные релаксаторы: аденозин, АМФ, АТФ при скарификации эндотелия повышают тонус гладких миоцитов.
Наличие этих различий, то есть способность в норме регулировать дилятацию изнутри, а констрикцию снаружи сформировано условиями развития кровеносных сосудов. Первичные сосуды закладываются как тонкостенные эндотелиальные трубки и кровь, замкнутая в них, является главным раздражителем их клеток. Давление ее действует в направлении расширения сосуда, то есть становится позиционной информацией, обеспечивающей формирование механизма релаксации. Согласно закону Ньютона всякое действие имеет равное, но противоположно направленное сопротивление. Им оказываются силы тканей, окружающих сосуд, которые стремятся удержать его в определенных размеров, образуя вокруг него, согласно гемодинамическим условиям, мышечную оболочку. Информация противодействия индуцирует развитие со стороны адвентиции механизмов, поддерживающих в мышце высокий тонус.
Таким образом, условия онтогенеза предопределяют различия в механизмах регуляции двигательного поведения кровеносных сосудов. Экспериментальные исследования подтверждают неравнозначность вазомоторных ответов при прямом и опосредованном через эндотелий действии на миоциты одного и того же вазоактивного фактора. Эти данные должны учитываться в клинической практике и могут быть эффективно использованы при фармакологической коррекции сосудистых нарушений в мозге.
Литература.
- Мотавкин П. А., Пиголкин Ю. И., Каминский Ю. В. Гистофизиология кровообращения в спинном мозге. — М.: Наука, 1994. – 233 с.
- Журнал "Морфология" 1996г. №2, стр.54.
- Журнал "Морфология" 1996г. №6, стр. 37.
- Тихоокеанский медицинский журнал 1998г. №1, стр.95.